See artikkel süveneb neuroplastilisuse ja selle sügava mõju uurimisse kognitiivsele kasvule. Artiklis pakutakse rakendatavaid viise neuroplastsuse ja aju kohanemisvõime suurendamiseks. Uurige kognitiivseid harjutusi, tähelepanelikkust ja meelte kaasamist, et edendada aju paindlikkust. Avastage seosed kehalise aktiivsuse, toitumise, une ja optimaalse neuroplastilisuse vahel.
Sissejuhatus
Neuroplastilisus, tuntud ka kui aju plastilisus või neuraalne plastilisus, viitab aju võimele end ümber korraldada, moodustades uusi närviühendusi ja muutes olemasolevaid. Seda võib nimetada ka protsessiks, mis hõlmab aju adaptiivseid struktuurilisi ja funktsionaalseid muutusi. Neuroplastilisus on muutnud meie arusaamist ajust, andes teadusliku aluse inimaju tähelepanuväärsele vastupidavusele ja kohanemisvõimele(1) .
Neuroplastilisuse idee pakkus esmakordselt välja 20. sajandi alguses Santiago Ramon y Cajal, kaasaegse neuroteaduse isa(2) . Kuid alles 20. sajandi teisel poolel saavutas see mõiste laialdase tunnustuse tänu neuroteaduslike uuringute ja pildistamistehnoloogia arengule.
Uuringud on näidanud, et aju muutub pidevalt vastuseks sisemistele ja välistele stiimulitele(3) . Iga kogemus, mõte ja emotsioon võib muuta meie aju struktuuri ja funktsiooni. Näiteks uue oskuse õppimine, näiteks muusikainstrumendi mängimine, žongleerimine või uue keele rääkimine, võib põhjustada uusi ühendusi neuronite vahel. Samal ajal võivad traumaatilised sündmused viia ühenduste kadumiseni.
Neuroplastilisuse aluseks olevad mehhanismid hõlmavad muutusi neuronite vaheliste ühenduste tugevuses ja arvus ning uute neuronite ja sünapside moodustumist. Neid muutusi juhib geneetiliste, epigeneetiliste ja keskkonnategurite, sealhulgas treeningu, toitumise, stressi ja sotsiaalse suhtluse keerukas koostoime(4) .
Neuroplastilisuse üks põnevamaid tagajärgi on see, et seda saab kasutada taastumise ja rehabilitatsiooni edendamiseks pärast vigastusi või haigusi. Näiteks insuldipatsientide puhul võib intensiivne taastusravi soodustada uute ühenduste kasvu ajus ja parandada motoorseid funktsioone. Samamoodi võib tähelepanelikkusel põhinev meditatsioon vähendada kroonilise valu all kannatavatel inimestel nende ajupiirkondade mahtu, mis töötlevad valusignaale.
Neuroplastilisuse eri liigid
Neuroplastilisuse võib laias laastus jagada kaheks peamiseks mehhanismiks: struktuurne plastilisus ja funktsionaalne plastilisus. Seoses indiviidi ajaskaalaga võib neuroplastilisuse jagada ka kahte faasi - arenguplastilisus ja täiskasvanu plastilisus. Need on kaks neuroplastilisuse aspekti, mis esinevad erinevates eluetappides.

Struktuurne plastilisus
Struktuurne plastilisus viitab füüsilistele muutustele ajus, näiteks sünapside moodustumisele või kadumisele, dendriitide okastraatide kasvule või tõmbumisele ning neuronite tekkimisele või kadumisele. Arvatakse, et need muutused on aluseks aju võimele kohaneda uute keskkondade ja kogemustega ning on eriti olulised arengu ajal, kui aju kasvab ja muutub kiiresti(5) .
Funktsionaalne plastilisus
Funktsionaalne plastilisusviitab seevastu muutustele neuronaalsete ahelate funktsionaalsetes omadustes, näiteks sünaptiliste ühenduste tugevuse muutustele või muutustele neuronivõrkude aktiivsusmustris. Need muutused on aluseks aju võimele õppida ja mäletada ning kohaneda muutuvate kognitiivsete nõudmiste ja keskkonnatingimustega(6) .
Struktuuriline ja funktsionaalne plastilisus on sageli üksteisest sõltuvad, kusjuures muutused ühes mehhanismis mõjutavad teist. Nii struktuurne kui ka funktsionaalne plastilisus on neuroplastilisuse kriitilised komponendid, mis võimaldavad ajul kohaneda ja muutuda vastuseks kogemustele ja keskkonnast tulenevatele stiimulitele(7) .
Arengu plastilisus
Arenguplastilisus ja täiskasvanute plastilisus on kaks neuroplastilisuse aspekti, mis toimuvad erinevates eluetappides. Arenguplastilisus viitab neuronaalse plastilisuse protsessile, mis toimub aju arengu ajal, alates embrüonaalsest arengust kuni lapsepõlve ja noorukieani. Selle aja jooksul on aju väga paindlik ja reageerib kogemustele, kusjuures neuronite ühendused ja ahelad kujunevad ja täiustuvad vastusena sensoorsetele sisenditele ja keskkonnast tulenevatele stiimulitele. Arenguplastilisus mängib aju normaalses arengus kriitilist rolli, sealhulgas funktsionaalsete närvikontuuride moodustamisel ja kriitiliste kognitiivsete ja käitumisfunktsioonide loomisel(8) .
Täiskasvanute plastilisus
Seevastu, täiskasvanu plastilisus viitab aju võimele teha täiskasvanueas läbi plastilisi muutusi vastuseks kogemustele või vigastustele. Kuigi plastilisuse tase on täiskasvanueas üldiselt madalam kui arengu ajal, on täiskasvanu aju siiski märkimisväärselt võimeline neuronaalseks plastilisuseks.
Täiskasvanute plastilisus on pidev õppimis- ja kohanemisprotsess, mis toimub kogu elu jooksul. See mängib olulist rolli kognitiivsete ja käitumisfunktsioonide säilitamisel muutuvas keskkonnas(9) .
Üks oluline erinevus arenguplastilisuse ja täiskasvanute plastilisuse vahel on plastiliste muutuste olemus. Arengu ajal hõlmab plastilisus sageli uute sünapside moodustamist, kasutamata ühenduste kärpimist ning dendriitiliste ja aksonaalsete protsesside kasvu ja ümberkorraldamist. Seevastu täiskasvanute plastilisus hõlmab olemasolevate ühenduste tugevdamist või nõrgenemist sünaptilise tugevuse muutuste kaudu ja uute ühenduste kasvu uute sünapside moodustamise või uute dendriitiliste protsesside idanemise kaudu.
Neuroplastilisuse põhikomponendid
1. Sünaptiline plastilisus
Sünaptiline plastilisus viitab sünapside ehk neuronite vaheliste ühenduste võimele muuta oma tugevust vastuseks aktiivsusele. See on õppimise ja mälu ning uute neuronühenduste moodustamise aluseks olev põhiline mehhanism. Sünaptiline plastilisus võib esineda nii erutavate kui ka pärssivate sünapside puhul ning seda põhjustavad muutused neurotransmitterite vabanemises ja retseptorite ekspressioonis postsünaptilisel membraanil.
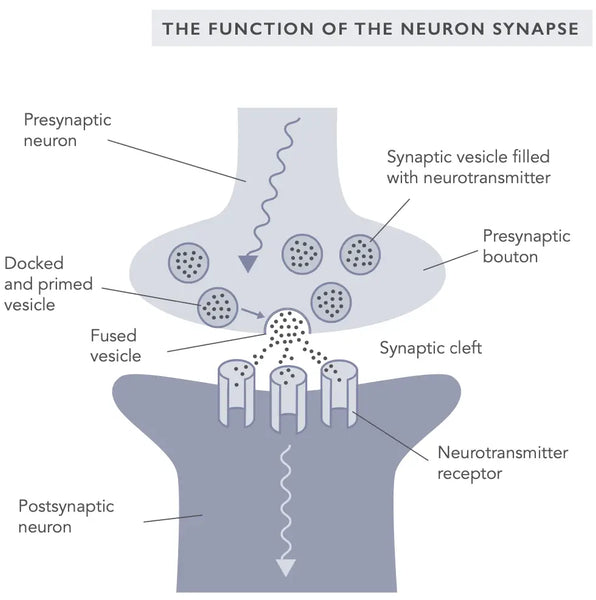
Kaks kõige paremini uuritud sünaptilise plastilisuse vormi on pikaajaline potentsiatsioon (LTP) ja pikaajaline depressioon (LTD), mida nimetatakse ka Hebbi plastilisuseks (viitab neuropsühholoog Donald Hebbile, kes tutvustas sünaptilist plastilisust esmakordselt 1949. aastal).(10)
LTP on protsess, mille käigus sünapsi tugevus suureneb vastuseks korduvale aktiivsusele. Arvatakse, et see on aluseks närviühenduste tugevnemisele õppimise ja mälu kujunemise ajal. LTD seevastu on protsess, mille puhul sünapsi tugevus väheneb vastuseks madala sagedusega või pikaajalise aktiivsuse korral. Arvatakse, et LTD mängib rolli neuronaalsete ühenduste nõrgenemisel unustamise ja kustutamise ajal(11) . Read rohkem LTP, LTD ja mälu kujunemise kohta alates Biohackeri käsiraamatu peatükist "Mind".
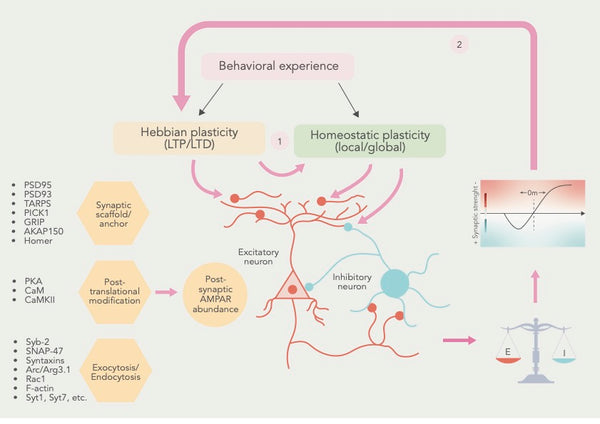
Joonis: Hebbiani ja homöostaatilise plastilisuse vaheline koostoimiv suhe.
Allikas: Li, J. & Park, E. & Zhong, L. & Chen, L. (2019). Homöostaatiline sünaptiline plastilisus kui metaplastilisuse mehhanism - molekulaarne ja rakuline perspektiiv. Current Opinion in Neurobiology 54: 44–53.
E= sünaptiline erutus I = sünaptiline inhibeerimine
Lisaks LTP-le ja LTD-le on tuvastatud palju teisi sünaptilise plastilisuse vorme, sealhulgas metaplastilisus, mis viitab LTP ja LTD indutseerimise lävendi muutustele ja homöostaatiline plastilisus, mis viitab neuronite võimele kohandada oma aktiivsust vastuseks võrgu aktiivsuse muutustele(12) .
Neid sünaptilise plastilisuse vorme kokku pannes on teadlased jõudnud järeldusele, et Hebbi ja homöostaatiline sünaptiline plastilisus lähenevad ühistele rakuprotsessidele ja et homöostaatiline plastilisus kohandab sünapside seisundit, et mõjutada Hebbi plastilisust (vt joonis eespool).
Sünaptilist plastilisust reguleerivad mitmesugused molekulaarsed ja rakulised mehhanismid, sealhulgas valgukinaaside ja fosfataaside aktiivsus, valkude süntees ja lagundamine ning muutused geeniekspressioonis. Need mehhanismid on tundlikud erinevate keskkonna- ja kogemustegurite suhtes, sealhulgas sensoorse sisendi, stressi ja sotsiaalse suhtluse suhtes(13-15).
Neurogeneesi
Neurogeneesi on protsess, mille käigus tekivad ajus uued neuronid, eriti hipokampuses, mis on oluline õppimise ja mälu jaoks. See on põhiline mehhanism, mis on aluseks aju võimele kohaneda ja reageerida keskkonna- ja kogemusteguritele.
Neurogeneesi toimub hipokampuse hambulise kihelkonna subgranulaarses tsoonis, kus neuronaalsed tüvirakud tekitavad vahepealseid progenitorrakke, millest omakorda tekivad ebaküpsed neuronid. Need ebaküpsed neuronid rändavad seejärel hipokampuse graanulite rakukihti, kus nad küpsevad ja integreeruvad olemasolevasse neuraalsesse ahelasse. Kuigi uusi neuroneid on avastatud ka teistes piirkondades, on neurogeneesi ulatus nendes piirkondades, näiteks neokorteksis ja hüpotalamuses, endiselt vastuoluline(16).
Neurogeneesi reguleerimine on keeruline ja dünaamiline protsess, mida mõjutavad erinevad tegurid, sealhulgas geneetika, epigeneetika ja keskkonnategurid, nagu treening ja stress. Näiteks on uuringud näidanud, et treening (eriti aeroobne treening) võib stimuleerida neurogeneesi, vabastades selliseid kasvufaktoreid nagu ajust saadav neurotroofiline faktor (BDNF) ja insuliinilaadne kasvufaktor-1 (IGF-1). Seevastu on näidatud, et stress ja krooniline põletik kahjustavad neurogeneesi põletikupõhise tsütokiini IL-1β kaudu (17-19).
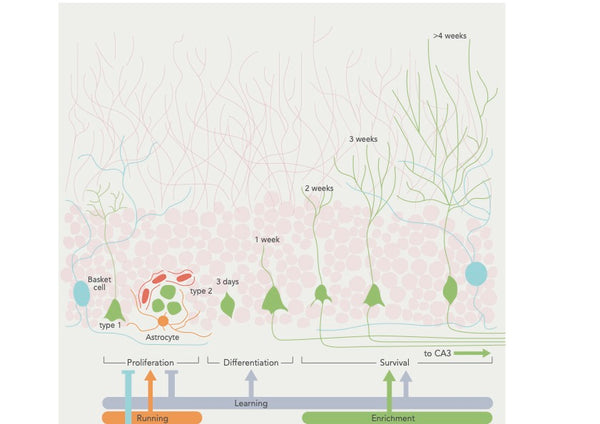
Joonis: Neurogeneesi reguleerimine käitumuslikest teguritest.
Allikas: Aimone, J. et al. (2014). Täiskasvanute neurogeneesi regulatsioon ja funktsioon: geenidest kognitsiooni. Physiological Reviews 94 (4): 991–1026.
Neurogeneesi funktsionaalne roll on endiselt aktiivse uurimistöö objektiks, kuid arvatakse, et see mängib rolli õppimises, mälus, meeleolu reguleerimises ja stressireaktsioonis. Neurogeneesi uurimine mõjutab oluliselt uute ravimeetodite ja sekkumiste väljatöötamist neuroloogiliste ja psühhiaatriliste seisundite puhul(20-22).
Dendriitide arboreerimine
Dendriitiline arborisatsioon (või dendriitiline hargnemine) viitab protsessile, mille käigus dendriidid, neuroni rakukehast välja ulatuvad hargnenud struktuurid, arenevad ja arendavad oma hargnemismustreid. See protsess on oluline aju neuronaalsete ahelate ühenduvuse ja funktsionaalsete omaduste loomiseks.
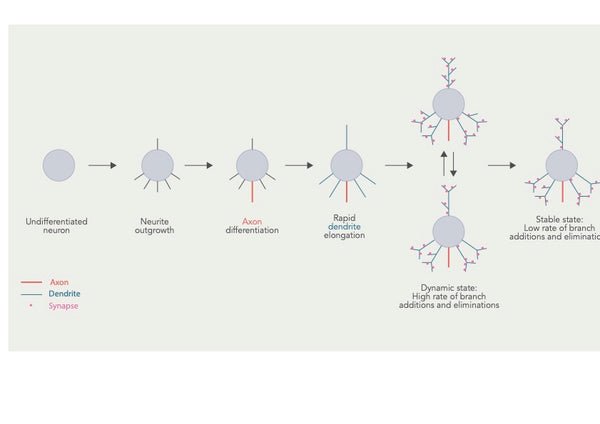 Joonis: Dendriitilise arbor'i areng ja mitmed kattuvad staadiumid.
Joonis: Dendriitilise arbor'i areng ja mitmed kattuvad staadiumid.
Allikas: Urbanska, M. & Blazejczyk, M. & Jaworski, J. (2008). Dendriitilise arborisatsiooni molekulaarsed alused. Acta Neurobiologiae Experimentalis 68 (2): 264–288.
Dendriitide arborisatsioon on keeruline protsess, mida reguleerivad erinevad tegurid, sealhulgas geneetilised ja epigeneetilised tegurid ning keskkonnategurid, näiteks sensoorne sisend ja neuraalne aktiivsus. Dendriitide kasvu ja hargnemist juhivad signaaliradade aktiivsus, mida aktiveerivad rakuvälised signaalid, näiteks kasvufaktorid ja neurotransmitterid. Need signaalid võivad mõjutada dendriitide kasvus ja hargnemises osalevate geenide ekspressiooni(23) .
Dendriitide arborisatsiooni reguleerimine on oluline funktsionaalsete neuraalsete ahelate loomiseks ja säilitamiseks ajus. Näiteks on dendriitide arborisatsioon kriitilise tähtsusega sünapside, neuronite vahelise kommunikatsiooni kohtade moodustamisel. Dendriitide hargnemismustrid võivad mõjutada moodustatud sünapside tüüpe ja arvu, mis võib oluliselt mõjutada neuronaalsete ahelate funktsionaalseid omadusi(24).
Dendriitide arborisatsioon mängib rolli sensoorse teabe töötlemisel. See on oluline ka kognitiivsete funktsioonide ja mälu moodustamisel. Täpsemalt öeldes võivad dendriitide hargnemismustrid mõjutada moodustatud sünapside tüüpe ja arvu, mis võib oluliselt mõjutada õppimises ja mälus osalevate neuraalsete ahelate funktsionaalseid omadusi(25).
Uuringud on näidanud, et dendriitide arborisatsiooni muutused võivad toimuda vastuseks õpikogemustele ning arvatakse, et need muutused aitavad kaasa uute mälestuste kujunemisele ja säilitamisele. Näiteks on näriliste ruumilise mälu ülesande treenimine suurendanud dendriitide hargnemist hipokampuses, mis on ruumilise õppimise ja mälu jaoks kriitiline ajupiirkond(26) .
Lisaks on uuringud näidanud, et dendriitide arborisatsiooni muutused on seotud neurodegeneratiivsete haiguste kognitiivsete puudujääkidega. Alzheimeri tõve korral kaovad dendriitilised selgrood - dendriitide struktuurid, mis moodustavad sünapsi teiste neuronitega - kahjustatud ajupiirkondades, mis põhjustab sünaptilise plastilisuse ja kognitiivsete puudujääkide kahjustumist(30) .
Müeliniseerumine
Müeliniseerumine on bioloogiline protsess, mille käigus aksonid, neuronite piklikud ja peenikesed rakulised pikendused, mis edastavad elektrilisi impulsse teistele neuronitele, ümbritsetakse lipiidirikka ainega, mida nimetatakse müeliiniks. Müeliinikile toodavad kesknärvisüsteemis (CNS) oligodendrotsüüdid ja perifeerses närvisüsteemis (PNS) Schwanni rakud. Müeliin toimib isolaatorina, võimaldades elektriliste signaalide kiiremat ja tõhusamat liikumist mööda aksoneid(27) .
Müeliniseerumisprotsess algab embrüonaalse arengu ajal ja jätkub varases täiskasvanueas, kusjuures eri aju ja närvisüsteemi piirkonnad müeliniseeruvad eri aegadel. Üldiselt algab müeliniseerumine ajutüves ja seljaajus ning liigub edasi ajukoorele ja teistele kõrgematele ajupiirkondadele(28). Müeliinikihid jäävad tavaliselt pikaks ajaks samapikkuseks, mis viitab sellele, et olemasoleva müeliini struktuur ei muutu oluliselt (vt allpool olevat pilti)(29).
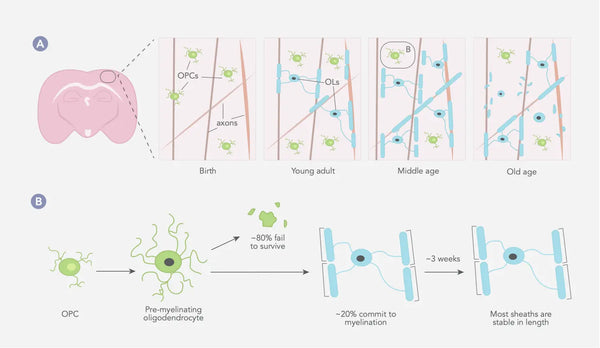
Joonis: Oligodendrotsüütide ja müeliini dünaamika imetajate somatosensoorses koores kogu elu jooksul.
Allikas: Williamson, J. & Lyons, D. (2018). Müeliini dünaamika kogu elu jooksul: pidevalt muutuv maastik? Frontiers in Cellular Neuroscience 12: 424.
OPC = Oligodendrotsüütide eellaste rakud. OL = Oligodendrotsüüdid
Müeliniseerumise regulatsioon on keeruline protsess, mida mõjutavad erinevad tegurid, sealhulgas geneetika, epigeneetika ja keskkonnategurid, näiteks kogemused ja närviaktiivsus. Näiteks on uuringud näidanud, et sensoorsed kogemused võivad mõjutada aju müeliniseerumise ajastust ja ulatust. Samamoodi võib neuronaalne aktiivsus soodustada müeliniseerumist, vabastades selliseid signaalmolekule nagu BDNF.
Kesknärvisüsteemis aktiveerivad müeliniseerumisprotsessi aksonaalne aktiivsus ja astrotsüüdid, samal ajal kui mikroglia/makrofaagid vastutavad müeliini eemaldamise eest. Kui aksonid on müeliniseerunud, sõltub nende jätkuv tervis ja funktsionaalsus oluliste metaboliitide ja neurotroofiliste faktorite pakkumisest gliarakkude poolt(31).
Müeliniseerumise funktsionaalne roll on kriitilise tähtsusega närvisignaalide tõhusaks ja tulemuslikuks edastamiseks ajus ja närvisüsteemis. Müeliniseerumine on võtmetähtsusega kognitiivsete ja motoorsete funktsioonide, sealhulgas tähelepanu, õppimise ja koordinatsiooni puhul. Lisaks sellele on müeliniseerumine vajalik ka valge aine, aju aksonühenduste võrgustiku arendamiseks, mis võimaldab erinevatel ajupiirkondadel suhelda ja koordineerida oma tegevust(32) .
Kortikaalne reorganiseerumine
Kortikaalne reorganiseerumine, mida nimetatakse ka kortikaalne plastilisusviitab aju võimele reorganiseerida oma närvivõrgustikke vastuseks muutustele sensoorse sisendi või muude kogemuste kujul. See protsess on kriitilise tähtsusega funktsionaalsete neuraalsete ahelate arengu ja aju võime jaoks kohaneda keskkonna muutustega. Kortikaalne reorganiseerumine toimub mitmel aju tasandil, alates primaarsetest sensoorsetest piirkondadest kuni kõrgema taseme assotsiatsioonipiirkondadeni(33).
Kortikaalse reorganiseerumise aluseks olevad mehhanismid hõlmavad muutusi sünaptilises tugevuses ja neuronite ühenduvuses. Näiteks on uuringud näidanud, et muutused sensoorses sisendmaterjalis võivad põhjustada muutusi sünapside võimsuses ja arvus mõjutatud kortikaalsetes piirkondades. Samamoodi võivad muutused kogemustes või käitumises põhjustada muutusi neuronaalse aktiivsuse mustris ning sünaptiliste ühenduste tugevuses ja spetsiifilisuses(34).
Kortikaalse reorganiseerimise funktsionaalsed tagajärjed võivad olla kasulikud või kahjulikud. Ühest küljest võib kortikaalne reorganiseerumine võimaldada ajul kohaneda keskkonnamuutustega ja taastuda vigastusest või haigusest. Teisest küljest võib kortikaalne reorganiseerumine aidata kaasa ka maladaptiivsete neuraalsete ahelate ja krooniliste valusündroomide tekkimisele.
Teine näide on tinnitusseisund, mille puhul inimesed tajuvad helinat või muud heli ilma välise stiimulita. Uuringud on näidanud, et tinnituse tekkimisel ja säilitamisel võib olla roll kuulmekoores toimuval kortikaalsel reorganiseerumisel. Konkreetselt võib aju reorganiseeruda vastusena kuulmisüsteemi kahjustusele, mis viib fantoomhelide tajumiseni(35) .
Kortikaalset reorganiseerimist soodustavad sekkumised (nt aju plastilisusel põhinevad ravimeetodid) võivad olla kasulikud krooniliste valusündroomide, insuldi ja muude neuroloogiliste kahjustuste ravimisel(36).
Looduslikud tegurid, mis suurendavad neuroplastilisust
On näidatud, et mitmed looduslikud ja tehnoloogilised meetodid soodustavad neuroplastilisust ja parandavad aju funktsiooni.
Allpool on loetletud parimad üldised elustiilifaktorid neuroplastilisuse parandamiseks:
- Uni: Piisav uni (ja sügav unieriti) on aju toimimise seisukohalt oluline ja on tõestatud, et see soodustab neuroplastilisust, suurendades sünaptilist plastilisust ja hõlbustades mälestuste konsolideerimist ning suurendades õppimisvõimet(37-38).(37-38)
- Harjutus: On näidatud, et füüsiline treening suurendab neuroplastilisust, soodustades uute neuronite moodustumist, suurendades dendriitide okastraatide kasvu ja parandades olemasolevate närvivõrgustike toimimist. Eelkõige on näidatud, et aeroobne treening suurendab neurotroofilisi faktorid (BDNF, NGF ja GDNF ), mis on valgud, mis soodustavad neuronite ja gliarakkude kasvu ja ellujäämist.(39-41)
- Meditatsioon: On näidatud, et mindfulness-meditatsioon soodustab neuroplastilisust, suurendades halli aine tihedust ajupiirkondades, mis on seotud tähelepanu, emotsioonide reguleerimise ja eneseteadvusega. Samuti võib see parandada valge aine terviklikkust, mis on oluline erinevate ajupiirkondade vaheliseks suhtlemiseks(42-44).
- Aeg-ajalt paastumine: Aeg-ajalt paastumine, mis hõlmab igapäevase toidu tarbimise piiramist teatud tundide jooksul, suurendab neuroplastilisust, soodustades uute neuronite kasvu ja sünaptilist plastilisust. Samuti võib see parandada kognitiivseid funktsioone ja vähendada neurodegeneratiivsete haiguste riski(45).
- Ajutine ainevahetuse vaheldumine (IMS): Eluviis, mis hõlmab vaheldumisi metaboolse stressi ja taastumise perioode, nagu paastumine ja treening, millele järgneb söömine, puhkus ja uni, on soovitanud parandada aju funktsiooni ja vastupidavust. IMS võib edendada kognitiivseid võimeid ja emotsionaalset heaolu toetavate neuronite tervist ja toimimist kogu elu jooksul. See mõjutab laias laastus mitmeid signaaliradu, mis suurendavad neuroplastilisust ja suurendavad aju vastupanuvõimet vigastuste ja haiguste vastu(46).
- Sotsiaalne kaasatus: On tõestatud, et sotsiaalne suhtlus ja kaasamine soodustavad neuroplastilisust, suurendades uute neuronite kasvu ja parandades olemasolevate neuronivõrgustike funktsiooni. Mõned uuringud on näidanud, et sotsiaalne kaasatus võib isegi kaitsta kognitiivse languse ja neurodegeneratiivsete haiguste, näiteks Alzheimeri tõve tekke eest. Seetõttu võib tugeva sotsiaalse võrgustiku säilitamine ja regulaarne sotsiaalne tegevus olla tõhus viis neuroplastilisuse toetamiseks ja suurendamiseks kogu elu jooksul(47-48).
- Keskkonna rikastamine: Lähenemisviis, mille puhul organismi elutingimused on optimeeritud, et pakkuda mitmekesist sensoorset, kognitiivset ja motoorset stimulatsiooni. On leitud, et see strateegia soodustab neuroplastilisust, indutseerides muutusi neuronaalses aktiivsuses ja morfoloogias. Konkreetselt on näidatud, et keskkonna rikastamine suurendab uute neuronite kasvu, soodustab sünaptilist plastilisust ja parandab olemasolevate neuronivõrgustike funktsiooni, mille tulemuseks on paremad kognitiivsed, käitumuslikud ja emotsionaalsed tulemused.(49-50)
- Kognitiivne koolitus: Aju neuroplastilisuse parandamiseks aju proovile panevad tegevused. Nende hulka kuuluvad uue keele õppimine, muusikainstrumendi mängimine või mõistatuste lahendamine - need võivad parandada neuroplastilisust, soodustades uute neuronite kasvu ja sünaptilist plastilisust.(51-52)
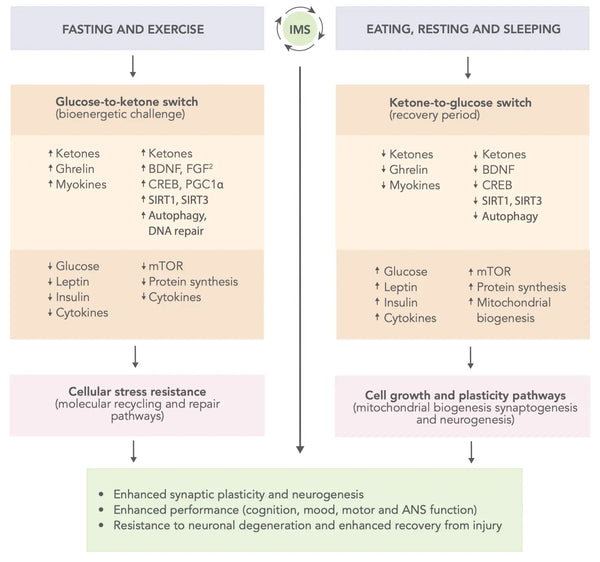
Joonis: Skeemiline mudel selle kohta, kuidas vahelduv ainevahetus võib optimeerida aju jõudlust ja suurendada vastupanu vigastustele ja haigustele.
Allikas: Mattson, M. & Moehl, K. & Ghena, N. & Schmaedick, M. & Cheng, A. (2018). Ajutine ainevahetuse ümberlülitamine, neuroplastilisus ja aju tervis. Nature Reviews Neuroscience 19 (2): 81–94.
Neuroplastilisust toetavad toitumistegurid
- Omega-3 rasvhapped: On tõestatud, et pika ahelaga oomega-3-rasvhapped, eriti dokosaheksaeenhape (DHA), soodustavad neuroplastilisust, suurendades sünaptilist plastilisust ja suurendades dendriitiliste selgroogade kasvu. Nad võivad vähendada ka põletikku ajus, mis võib kahjustada neuroplastilisust. Oomega-3-rasvhapete allikad on rasvased kalad, nagu lõhe ja sardiinid, ning toidulisandid(53-55).
- Kurkumiin: Kurkumiin, kurkumas leiduv ühend, on näidanud, et see suurendab neuroplastilisust, soodustades uute neuronite kasvu ja tugevdades sünaptilist plastilisust. Sellel võib olla ka põletikuvastane toime, mis võib parandada aju toimimist. Huvitaval kombel võib kurkumiin ka kroonilise stressi poolt põhjustatud kognitiivsete võimete ja neuronaalse plastilisuse kahjustusi tagasi pöörata(56-57). Proovige AGEless Defense toidulisandit, mis sisaldab B-vitamiine, polüfenoole, aminohappeid ja kurkumiini.
- B-vitamiinid: On tõestatud, et B-kompleksi vitamiinid ja koliin parandavad aju neuroplastilisust. Neil on oluline roll erinevates ainevahetusradades, mis toetavad aju toimimist, sealhulgas neurotransmitterite ja müeliini sünteesimisel. B-vitamiinid, eriti vitamiin B12 ja folaat, osalevad ka DNA metülatsioonis, mis võib mõjutada geeniekspressiooni neuroplastilisuses(58-62).
- D-vitamiin: Mõned tõendid viitavad sellele, et D-vitamiin võib mängida rolli neuroplastilisuse edendamisel. D-vitamiini retseptoreid on leitud erinevates ajupiirkondades, sealhulgas hipokampuses, mis on seotud õppimise ja mäluga. Loomadel ja inimestel tehtud uuringud on näidanud, et D-vitamiini puudus võib kahjustada kognitiivseid funktsioone ja vähendada neuroplastilisuse edendamiseks vajalike spetsiifiliste neurotroofsete faktorite tootmist(63-64) . Hankige Ecosh K2+D3-vitamiini, et maksimeerida neuroplastilisust.
- Polüfenoolid: On tõestatud, et polüfenoolid parandavad aju neuroplastilisust. Üks mehhanismidest on nende võime moduleerida sünaptilises plastilisuses ja neurogeneesis osalevaid signaaliradu. Samuti võivad nad avaldada põletikuvastast mõju ja kaitsta oksüdatiivse stressi eest, parandades neuronite funktsiooni ja edendades neuroplastilisust. Üldiselt võivad polüfenoolid ennetada neurodegeneratiivsete patoloogiate progresseerumist. 65-66 - Proovi üks parimaid polüfenoole: Purovitalis Liposomal Quercetin

Neuroplastilisust toetavad tehnoloogilised meetodid
- transkraniaalne magnetiline stimulatsioon (TMS): TMS on mitteinvasiivne tehnika, mis kasutab magnetvälju, et stimuleerida neuraalset aktiivsust konkreetsetes ajupiirkondades. On näidatud, et see suurendab neuroplastilisust ja parandab kognitiivseid funktsioone erinevates kontekstides, sealhulgas depressiooni, ärevuse ja insuldiga inimestel(67-69) . Hankige NeoRhythm OmniPEMF neurostimulatsiooniseade siit. [vt pilti allpool]
- Aju stimuleerimine: Lisaks TMS-ile on näidatud, et teised ajustimulatsiooni vormid, nagu transkraniaalne alalisvoolustimulatsioon (tDCS) ja transkraniaalne vahelduvvoolustimulatsioon (tACS), suurendavad neuroplastilisust ja parandavad kognitiivseid funktsioone(70-71).(70-71) Lisateavet tDCS-i kohta leiate veebilehelt Biohackeri käsiraamatust.
- Neurofeedback: Neurofeedback on tehnika, mis hõlmab aju elektrilise aktiivsuse jälgimist ja tagasiside andmist inimesele reaalajas. On tõestatud, et see suurendab neuroplastilisust, soodustades uute neuronite kasvu ja sünaptilist plastilisust. Neurofeedbacki on kasutatud tähelepanu puudulikkuse ja hüperaktiivsuse häire (ADHD), ärevuse ja teiste neuroloogiliste seisundite raviks.(72-74) Lugege rohkem neurofeedbacki kohta järgmiselt Biohackeri käsiraamatust.
- Virtuaalne reaalsus (VR): Virtuaalreaalsus on immersiivne tehnoloogia koolituseks, rehabilitatsiooniks ja terapeutilistel eesmärkidel. Viimastel aastatel on suurenenud huvi VR-i kasutamise vastu aju neuroplastilisuse suurendamiseks. VR võib parandada neuroplastilisust, pakkudes stimuleerivat ja kaasahaaravat keskkonda, mis paneb aju kohanema ja õppima. Näiteks võib VR simuleerida tegeliku elu stsenaariume ning pakkuda võimalusi õppimiseks ja harjutamiseks turvalises ja kontrollitud keskkonnas. See võib aidata edendada uute neuronite kasvu ja suurendada sünaptilist plastilisust.(75-76)
- Kognitiivne koolitustarkvara: Kognitiivse treeningu tarkvara kasutab arvutiprogramme, et parandada kognitiivseid funktsioone, esitades ajule väljakutseid mäluharjutuste, probleemilahenduse ülesannete ja tähelepanuülesannete abil. Lisaks võib vaimne treening stimuleerida teatud neurotransmitterite, näiteks dopamiini ja atsetüülkoliini vabanemist, mis võib parandada sünaptilist plastilisust ja kognitiivset funktsiooni ning suurendada BDNF-i ajus(77-79).(77-79)

Järeldus
Kokkuvõtteks võib öelda, et aju tähelepanuväärne võime ennast ümber kujundada erinevate neuroplastilisuse vormide kaudu - arengu-, täiskasvanud, struktuuriline ja funktsionaalne - avab maailma võimalusi isikliku kasvu ja kognitiivse täiustamise jaoks. Selles artiklis esitatud strateegiate rakendamine annab teile vahendid, et alustada ümberkujundavat teekonda. Te kujundate aktiivselt oma aju tulevikku, tegeledes vaimsete harjutustega, kultiveerides tähelepanelikkust ja seades prioriteediks tervislikud harjumused. Pidage meeles, et võti peitub järjepidevuses ja pühendumuses. Niisiis, võtke vastu see põnev seiklus, et avada oma ajupotentsiaal.
Kui soovite oma mängu järgmisele oktaavile viia, tellige ette meie mahukas jätk raamat "Biohäkkerite käsiraamat", mis ongi Resilientse olemise raamatu.
P.S. See artikkel põhineb Resilientse olemise raamatu vaimse vastupidavuse osa tekstil.
Viited:
- Puderbaugh, M. & Emmady, P. (2022). Neuroplastilisus. In . StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing.
- Venkataramani, P. (2010). Santiago Ramón y Cajal: neuroteaduste isa. Resonants 15 (11): 968–976.
- Draganski, B. & Gaser, C. & Busch, V. & Schuierer, G. & Bogdahn, U. & May, A. (2004). Treeningu poolt esilekutsutud muutused hallis aines. Nature 427 (6972): 311–312.
- Kleim, J. & Jones, T. (2008). Kogemusest sõltuva neuraalse plastilisuse põhimõtted: mõju taastusravile pärast ajukahjustust. Journal of Speech Language and Hearing Research 51: S225-S239.
- Bozelos, P. & Poirazi, P. (2017). Struktuurse plastilisuse mõju mälu võimekusele. In . The Rewiring Brain (lk 319-341). Cambridge (USA): Academic Press.
- Grafman, J. (2000). Funktsionaalse neuroplastilisuse kontseptualiseerimine. Journal of Communication Disorders 33 (4): 345–356.
- Taubert, M. et al. (2010). Inimese aju struktuuri dünaamilised omadused: õppimisega seotud muutused kortikaalsetes piirkondades ja nendega seotud kiudühendustes. Journal of Neuroscience 30 (35): 11670–11677.
- Kolb, B. & Gibb, R. (2011). Aju plastilisus ja käitumine arenevas ajus. Journal of the Canadian Academy of Child and Adolescent Psychiatry (Kanada laste ja noorukite psühhiaatria akadeemia). 20 (4): 265–276.
- Fuchs, E. & Flügge, G. (2014). Täiskasvanute neuroplastilisus: rohkem kui 40 aastat uuringuid. Neural Plasticity 2014: 541870
- Hebb, D. (1949). Käitumise korraldus: Neuropsühholoogiline teooria. New York: John Wiley and Sons.
- Bliss, T. & Collingridge, G. (1993). Mälu sünaptiline mudel: pikaajaline potentsiatsioon hipokampuses. Nature 361 (6407): 31–39.
- Li, J. & Park, E. & Zhong, L. & Chen, L. (2019). Homöostaatiline sünaptiline plastilisus kui metaplastilisuse mehhanism - molekulaarne ja rakuline perspektiiv. Current Opinion in Neurobiology 54: 44–53.
- Magee, J. & Grienberger, C. (2020). Sünaptilise plastilisuse vormid ja funktsioonid. Annual Review of Neuroscience 43: 95–117.
- Vitureira, N. & De Pasquale, R. & Leão, R. & Rossi, F. (2022). Sünaptilise plastilisuse rakulised ja molekulaarsed mehhanismid hipokampuse ja kortikaalsete sünapside juures. Frontiers in Cellular Neuroscience 16: 980623.
- Fox, K. & Stryker, M. (2017). Hebbiani ja homöostaatilise plastilisuse integreerimine: sissejuhatus. Philosophical Transactions of the Royal Society B: Biological Sciences. 372 (1715): 20160413.
- Aimone, J. et al. (2014). Täiskasvanute neurogeneesi regulatsioon ja funktsioon: geenidest kognitsioonini. Physiological Reviews 94 (4): 991–1026
- Cotman, C. & Berchtold, N. (2002). Harjutus: käitumuslik sekkumine aju tervise ja plastilisuse parandamiseks. Trends in Neurosciences 25 (6): 295–301.
- Vecchio, L. et al. (2018). Treeningu neuroprotektiivne mõju: terve aju säilitamine kogu vananemise ajal. Brain Plasticity 4 (1): 17–52.
-
Saxe, M. et al. (2006). Hippokampuse neurogeneesi ablatsioon kahjustab kontekstuaalset hirmu konditsioneerimist ja sünaptilist plastilisust hambumuse gürosis. Proceedings of the National Academy of Sciences 103 (46): 17501–17506.
- Aimone, J. et al. (2014). Täiskasvanute neurogeneesi regulatsioon ja funktsioon: geenidest kognitsiooni. Physiological Reviews 94 (4): 991–1026.
- Ming, G. & Song, H. (2011). Täiskasvanud neurogeneesi imetajate ajus: olulised vastused ja olulised küsimused. Neuron . 70 (4): 687–702.
- Urbanska, M. & Blazejczyk, M. & Jaworski, J. (2008). Dendriitilise arborisatsiooni molekulaarsed alused. Acta Neurobiologiae Experimentalis 68 (2): 264–288.
- Cline, H. (2001). Dendriidi arborite areng ja sünaptogenees. Current Opinion In Neurobiology 11 (1): 118–126.
- Kasai, H. & Fukuda, M. & Watanabe, S. & Hayashi-Takagi, A. & Noguchi, J. (2010). Dendriitiliste selgroogade struktuuriline dünaamika mälu ja tunnetuse puhul. Trends in Neurosciences 33 (3): 121–129.
- Holtmaat, A. & Svoboda, K. (2009). Kogemusest sõltuv struktuurne sünaptiline plastilisus imetajate ajus. Nature Reviews Neuroscience 10 (9): 647–658.
- Demerens, C. et al. (1996). Müeliniseerumise indutseerimine kesknärvisüsteemis elektrilise aktiivsuse abil. Proceedings of the National Academy of Sciences 93 (18): 9887–9892.
- Harry, G. & Toews, A. (1998). Müeliniseerumine, düsmüeliniseerumine ja demüeliniseerumine. Handbook of Developmental Neurotoxicology (Arengu neurotoksikoloogia käsiraamat). 87–115.
- Williamson, J. & Lyons, D. (2018). Müeliini dünaamika kogu elu jooksul: pidevalt muutuv maastik? Frontiers in Cellular Neuroscience 12: 424.
- Scheff, S. & Price, D. & Schmitt, F. & Mufson, E. (2006). Hippokampuse sünaptiline kadu varajase Alzheimeri tõve ja kerge kognitiivse kahjustuse korral. Neurobiology of Aging 27 (10): 1372–1384.
- Nave, K. & Werner, H. (2014). Närvisüsteemi müeliniseerumine: mehhanismid ja funktsioonid. Aastakokkuvõte raku- ja arengubioloogia kohta 30: 503–533
- Fields, R. (2015). Uus närvisüsteemi plastilisuse mehhanism: aktiivsusest sõltuv müeliniseerumine. Nature Reviews Neuroscience 16 (12): 756–767.
- Pascual-Leone, A. & Amedi, A. & Fregni, F. & Merabet, L. (2005). Inimese plastiline ajukoor. Annual Reviews Neuroscience 28: 377–401.
- Schoups, A. & Vogels, R. & Qian, N. & Orban, G. (2001). Orienteerumise tuvastamise harjutamine parandab orientatsiooni kodeerimist V1 neuronites. Nature 412 (6846): 549–553.
- Mühlnickel, W. & Elbert, T. & Taub, E. & Flor, H. (1998). Kuulmiskoore reorganiseerumine tinnituse korral. Proceedings of the National Academy of Sciences 95 (17): 10340–10343.
- Merzenich, M. & Van Vleet, T. & Nahum, M. (2014). Aju plastilisusel põhinev ravi. Frontiers in Human Neuroscience 8: 385.
- Fattinger, S. et al. (2017). Sügav uni säilitab inimese aju õppimise tõhusust. Nature Communications 8 (1): 15405.
- Voss, M. et al. (2010). Aravavõrgustike plastilisus eakate täiskasvanute treeningu randomiseeritud sekkumisuuringus. Frontiers in Aging Neuroscience 2: 32.
- e Sousa Fernandes, M. et al. (2020). Füüsilise treeningu mõju neuroplastilisusele ja aju funktsioonile: süstemaatiline ülevaade inim- ja loomkatsetest. Neural Plasticity 2020: 8856621
- Hölzel, B. et al. (2011). Mindfulnessi praktika viib aju piirkondliku halli aine tiheduse suurenemiseni. Psühhiaatriauuringud: Neuroimaging 191 (1): 36–43.
- Tang, Y. & Hölzel, B. K. & Posner, M. (2015). Meeleolumeditatsiooni neuroteadus. Nature Reviews Neuroscience 16 (4): 213–225.
- Lardone, A. et al. (2018). Mindfulness-meditatsioon on seotud pikaajaliste muutustega hipokampuse funktsionaalses topoloogias puhkeoleku ajal: magnetoentsefalograafia uuring. Neural Plasticity 2018: 5340717.
- Brocchi, A. & Rebelos, E. & Dardano, A. & Mantuano, M. & Daniele, G. (2022). Ajutise paastumise mõju aju ainevahetusele. Nutrients 14 (6): 1275.
- Mattson, M. & Moehl, K. & Ghena, N. & Schmaedick, M. & Cheng, A. (2018). Ajutine ainevahetuse ümberlülitamine, neuroplastilisus ja aju tervis. Nature Reviews Neuroscience 19 (2): 81–94.
- Kramer, A. & Bherer, L. & Colcombe, S. & Dong, W. & Greenough, W. (2004). Keskkonna mõju kognitiivsele ja aju plastilisusele vananemise ajal. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences. 59 (9): M940-M957.
- Fratiglioni, L. & Paillard-Borg, S. & Winblad, B. (2004). Aktiivne ja sotsiaalselt integreeritud eluviis hilises elus võib kaitsta dementsuse eest. The Lancet Neurology 3 (6): 343–353.
- Kempermann, G. (2015). Aktiivsussõltuvus ja vananemine täiskasvanute neurogeneesi regulatsioonis. Cold Spring Harbor Perspectives in Biology 7 (11): a018929.
- Van Praag, H. & Kempermann, G. & Gage, F. (2000). Keskkonna rikastamise närvilised tagajärjed. Nature Reviews Neuroscience 1 (3): 191–198
- Lövdén, M. & Bäckman, L. & Lindenberger, U. & Schaefer, S. & Schmiedek, F. (2010). Teoreetiline raamistik täiskasvanute kognitiivse plastilisuse uurimiseks. Psychological Bulletin 136 (4): 659–676
- Park, D. & Bischof, G. (2013). Vananev mõistus: neuroplastilisus vastuseks kognitiivsele treeningule. Dialogues in Clinical Neuroscience 15 (1): 109–119.
- Crupi, R. & Marino, A. & Cuzzocrea, S. (2013). n-3 rasvhapped: roll neurogeneesis ja neuroplastilisuses. Current Medicinal Chemistry 20 (24): 2953–2963.
- Swanson, D. & Block, R. & Mousa, S. A. (2012). Omega-3-rasvhapped EPA ja DHA: kasu tervisele kogu elu jooksul. Advances in Nutrition 3 (1): 1–7.
- Dyall, S. (2015). Pikaahelalised oomega-3-rasvhapped ja aju: ülevaade EPA, DPA ja DHA sõltumatutest ja ühistest mõjudest. Frontiers in Aging Neuroscience 7: 52.
- Maharjan, R. et al. (2020). Elustiili roll neuroplastilisuses ja neurogeneesis vananevas ajus. Cureus 12 (9): e10639.
- Xu, Y. et al. (2009). Kurkumiin tühistab kroonilise stressi poolt esilekutsutud kognitsiooni ja neuronaalse plastilisuse häireid. Neuropharmacology 57 (4): 463–471.
- Echeverry, M. et al. (2021). Vitamiinid D ja B 12, muutunud sünaptiline plastilisus ja ekstratsellulaarne maatriks. In B-kompleksi vitamiinide allikad, tarbimine ja uued rakendused. IntechOpen.
- Downey, L. et al. (2019). Suurenenud tagumise tsingulaadi funktsionaalne ühenduvus pärast 6-kuulist suure annusega B-vitamiini multivitamiini lisamist: randomiseeritud, topeltpime, platseebokontrollitud uuring. Frontiers in Nutrition 6: 156.
- Mattson, M. & Shea, T. (2003). Folaadi ja homotsüsteiini ainevahetus neuraalses plastilisuses ja neurodegeneratiivsetes häiretes. Trends in Neurosciences 26 (3): 137–146.
- Chin, E. & Goh, E. (2019). Neuronaalse plastilisuse moduleerimine koliiniga. Neural Regeneration Research 14 (10): 1697.
- Jadavji, N. & Emmerson, J. & MacFarlane, A. & Willmore, W. & Smith, P. (2017). B-vitamiinide ja koliini lisamine suurendab neuroplastilisust ja taastumist pärast insulti. Neurobiology of Disease 103: 89–100.
- Mayne, P. & Burne, T. (2019). D-vitamiin sünaptilises plastilisuses, kognitiivsetes funktsioonides ja neuropsühhiaatrilistes haigustes. Trends in Neurosciences 42 (4): 293–306.
- Echeverry, M. et al. (2021). Vitamiinid D ja B 12, muutunud sünaptiline plastilisus ja ekstratsellulaarne maatriks. In B-kompleksi vitamiinide allikad, tarbimine ja uued rakendused. IntechOpen.
- Vauzour, D. (2012). Toidu polüfenoolid kui ajufunktsioonide modulaatorid: bioloogilised toimed ja molekulaarsed mehhanismid, mis toetavad nende kasulikku mõju. Oksüdatiivne meditsiin ja rakuline pikaealisus. 2012: 914273.
- Figueira, I. & Menezes, R. & Macedo, D. & Costa, I. & Nunes dos Santos, C. (2017). Polüfenoolid väljaspool tõkkeid: pilguheit ajju. Current Neuropharmacology 15 (4): 562–594.
- Hallett, M. (2007). Transkraniaalne magnetstimulatsioon: algkursus. Neuron 55 (2): 187–199.
- Jannati, A. & Oberman, L. & Rotenberg, A. & Pascual-Leone, A. (2023). Aju plastilisuse mehhanismide hindamine transkraniaalse magnetstimulatsiooni abil. Neuropsühhofarmakoloogia 48 (1): 191–208.
- Auriat, A. & Neva, J. & Peters, S. & Ferris, J. & Boyd, L. (2015). Ülevaade transkraniaalse magnetstimulatsiooni ja multimodaalse neuropildistamise kohta insuldijärgse neuroplastilisuse iseloomustamiseks. Frontiers in Neurology 6: 226.
- Kricheldorff, J. et al. (2022). Tõendid neuroplastiliste muutuste kohta pärast transkraniaalset magnetilist, elektrilist ja sügavat ajustimulatsiooni. Brain Sciences 12 (7): 929.
- Brunoni, A. et al. (2012). Kliinilised uuringud transkraniaalse otsevoolustimulatsiooniga (tDCS): väljakutsed ja tulevased suunad. Aju stimuleerimine 5 (3): 175–195.
- Gruzelier, J. (2014). EEG-neurofeedback jõudluse optimeerimiseks. III: ülevaade metoodilistest ja teoreetilistest kaalutlustest. Neuroscience & Biobehavioral Reviews. 44: 159–182.
- Trambaiolli, L. & Cassani, R. & Mehler, D. & Falk, T. (2021). Neurofeedback ja vananev aju: dementsuse ja kerge kognitiivse kahjustuse treeningprotokollide süstemaatiline ülevaade. Frontiers in Aging Neuroscience 13: 682683.
- Sitaram, R. et al. (2017). Suletud aju treening: neurofeedbacki teadus. Nature Reviews Neuroscience 18 (2): 86–100.
- Laver, K. & George, S. & Thomas, S. & Deutsch, J. & Crotty, M. (2015). Virtuaalreaalsus insuldi taastusravi jaoks: Cochrane'i ülevaate lühendatud versioon. European Journal of Physical and Rehabilitation Medicine 51 (4): 497–506.
- Huang, C. et al. (2022). Virtuaalreaalsusel põhineva motoorse kontrolli treeningu mõju põletikule, oksüdatiivsele stressile, neuroplastilisusele ja ülemiste jäsemete motoorse funktsiooni paranemisele kroonilise insuldiga patsientidel: randomiseeritud kontrollitud uuring. BMC Neurology 22 (1): 21.
- Anguera, J. et al. (2013). Videomängude treening parandab kognitiivset kontrolli eakatel täiskasvanutel. Nature 501 (7465): 97–101.
- Lampit, A. et al. (2014). Üldise kognitiivse kasu ajaline kulg juhendatud arvuti abil toimuva kognitiivse treeningu puhul: randomiseeritud, aktiivselt kontrollitud uuring mitme dementsuse riskiteguriga eakatel. The Journal of Prevention of Alzheimers Disease 1 (1): 33–39.
- D'Antonio, J. et al. (2019). Kognitiivne treening ja neuroplastilisus kerge kognitiivse kahjustuse korral (COG-IT): kahes kohas toimuva pimendatud, randomiseeritud, kontrollitud ravikatsetuse protokoll. BMJ Open 9 (8): e028536.